
Silurian
| Silurian | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
.png) A map of Earth as it appeared 430 million years ago during the Silurian Period, Wenlock Epoch | |||||||||||||
| Chronology | |||||||||||||
| |||||||||||||
| Etymology | |||||||||||||
| Name formality | Formal | ||||||||||||
| Synonym(s) | Gotlandian | ||||||||||||
| Usage information | |||||||||||||
| Celestial body | Earth | ||||||||||||
| Regional usage | Global (ICS) | ||||||||||||
| Time scale(s) used | ICS Time Scale | ||||||||||||
| Definition | |||||||||||||
| Chronological unit | Period | ||||||||||||
| Stratigraphic unit | System | ||||||||||||
| First proposed by | Roderick Murchison, 1835 | ||||||||||||
| Time span formality | Formal | ||||||||||||
| Lower boundary definition | FAD of the Graptolite Akidograptus ascensus | ||||||||||||
| Lower boundary GSSP | Dob's Linn, Moffat, United Kingdom 55°26′24″N 3°16′12″W / 55.4400°N 3.2700°W | ||||||||||||
| Lower GSSP ratified | 1984[4][5] | ||||||||||||
| Upper boundary definition | FAD of the Graptolite Monograptus uniformis | ||||||||||||
| Upper boundary GSSP | Klonk, Czech Republic 49°51′18″N 13°47′31″E / 49.8550°N 13.7920°E | ||||||||||||
| Upper GSSP ratified | 1972[6] | ||||||||||||
| Atmospheric and climatic data | |||||||||||||
| Sea level above present day | Around 180 m, with short-term negative excursions[7] | ||||||||||||
The Silurian (/sɪˈljʊəri.ən, saɪ-/ sih-LURE-ee-ən, sy-)[8][9][10] is a geologic period and system spanning 24.6 million years from the end of the Ordovician Period, at 443.8 million years ago (Mya), to the beginning of the Devonian Period, 419.2 Mya.[11] The Silurian is the third and shortest period of the Paleozoic Era, and the third of twelve periods of the Phanerozoic Eon. As with other geologic periods, the rock beds that define the period's start and end are well identified, but the exact dates are uncertain by a few million years. The base of the Silurian is set at a series of major Ordovician–Silurian extinction events when up to 60% of marine genera were wiped out.
One important event in this period was the initial establishment of terrestrial life in what is known as the Silurian-Devonian Terrestrial Revolution: vascular plants emerged from more primitive land plants,[12][13] dikaryan fungi started expanding and diversifying along with glomeromycotan fungi,[14] and three groups of arthropods (myriapods, arachnids and hexapods) became fully terrestrialized.[15]
Another significant evolutionary milestone during the Silurian was the diversification of jawed fish, which include placoderms, acanthodians (which gave rise to cartilaginous fish) and osteichthyan (bony fish, further divided into lobe-finned and ray-finned fishes),[16] although this corresponded to sharp decline of jawless fish such as conodonts and ostracoderms.
History of study
[edit]−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
The Silurian system was first identified by the Scottish geologist Roderick Murchison, who was examining fossil-bearing sedimentary rock strata in south Wales in the early 1830s. He named the sequences for a Celtic tribe of Wales, the Silures, inspired by his friend Adam Sedgwick, who had named the period of his study the Cambrian, from a Latin name for Wales.[17] Whilst the British rocks now identified as belonging to the Silurian System and the lands now thought to have been inhabited in antiquity by the Silures show little correlation (cf. Geologic map of Wales, Map of pre-Roman tribes of Wales), Murchison conjectured that their territory included Caer Caradoc and Wenlock Edge exposures - and that if it did not there were plenty of Silurian rocks elsewhere 'to sanction the name proposed'.[18] In 1835 the two men presented a joint paper, under the title On the Silurian and Cambrian Systems, Exhibiting the Order in which the Older Sedimentary Strata Succeed each other in England and Wales, which was the germ of the modern geological time scale.[19] As it was first identified, the "Silurian" series when traced farther afield quickly came to overlap Sedgwick's "Cambrian" sequence, however, provoking furious disagreements that ended the friendship.
The English geologist Charles Lapworth resolved the conflict by defining a new Ordovician system including the contested beds.[20] An alternative name for the Silurian was "Gotlandian" after the strata of the Baltic island of Gotland.[21]
The French geologist Joachim Barrande, building on Murchison's work, used the term Silurian in a more comprehensive sense than was justified by subsequent knowledge. He divided the Silurian rocks of Bohemia into eight stages.[22] His interpretation was questioned in 1854 by Edward Forbes,[23] and the later stages of Barrande; F, G and H have since been shown to be Devonian. Despite these modifications in the original groupings of the strata, it is recognized that Barrande established Bohemia as a classic ground for the study of the earliest Silurian fossils.
Subdivisions
[edit]| Epoch | Age | Start (mya) |
Etymology of Epochs and Stages |
Notes |
|---|---|---|---|---|
| Llandovery | Rhuddanian | 443.8 | Cefn-Rhuddan Farm, Llandovery in Carmarthenshire, Wales | |
| Aeronian | 440.8 | Cwm Coed-Aeron Farm, Wales | Trefawr Track near the farm is the site of the GSSP | |
| Telychian | 438.5 | Pen-lan-Telych Farm, Llandovery, Wales | ||
| Wenlock | Sheinwoodian | 433.4 | Sheinwood village, Much Wenlock and Wenlock Edge, Shropshire, England | During the Wenlock, the oldest-known tracheophytes of the genus Cooksonia, appear. The complexity of slightly later Gondwana plants like Baragwanathia, which resembled a modern clubmoss, indicates a much longer history for vascular plants, extending into the early Silurian or even Ordovician.[citation needed] The first terrestrial animals also appear in the Wenlock, represented by air-breathing millipedes from Scotland.[24] |
| Homerian | 430.5 | Homer, Shropshire, England | ||
| Ludlow | Gorstian | 427.4 | Gorsty village near Ludlow, Shropshire, England | |
| Ludfordian | 425.6 | Ludford, Shropshire, England | ||
| Přídolí | — | 423.0 | Named after a locality at the Homolka a Přídolí nature reserve near the Prague suburb of Slivenec, Czech Republic. | Přídolí is the old name of a cadastral field area.[25] |
Paleogeography
[edit]
With the supercontinent Gondwana covering the equator and much of the southern hemisphere, a large ocean occupied most of the northern half of the globe.[26] The high sea levels of the Silurian and the relatively flat land (with few significant mountain belts) resulted in a number of island chains, and thus a rich diversity of environmental settings.[26]
During the Silurian, Gondwana continued a slow southward drift to high southern latitudes, but there is evidence that the Silurian icecaps were less extensive than those of the late-Ordovician glaciation. The southern continents remained united during this period. The melting of icecaps and glaciers contributed to a rise in sea level, recognizable from the fact that Silurian sediments overlie eroded Ordovician sediments, forming an unconformity. The continents of Avalonia, Baltica, and Laurentia drifted together near the equator, starting the formation of a second supercontinent known as Euramerica.
When the proto-Europe collided with North America, the collision folded coastal sediments that had been accumulating since the Cambrian off the east coast of North America and the west coast of Europe. This event is the Caledonian orogeny, a spate of mountain building that stretched from New York State through conjoined Europe and Greenland to Norway. At the end of the Silurian, sea levels dropped again, leaving telltale basins of evaporites extending from Michigan to West Virginia, and the new mountain ranges were rapidly eroded. The Teays River, flowing into the shallow mid-continental sea, eroded Ordovician Period strata, forming deposits of Silurian strata in northern Ohio and Indiana.
The vast ocean of Panthalassa covered most of the northern hemisphere. Other minor oceans include two phases of the Tethys, the Proto-Tethys and Paleo-Tethys, the Rheic Ocean, the Iapetus Ocean (a narrow seaway between Avalonia and Laurentia), and the newly formed Ural Ocean.

Climate and sea level
[edit]The Silurian period was once believed to have enjoyed relatively stable and warm temperatures, in contrast with the extreme glaciations of the Ordovician before it and the extreme heat of the ensuing Devonian; however, it is now known that the global climate underwent many drastic fluctuations throughout the Silurian,[27][28] evidenced by numerous major carbon and oxygen isotope excursions during this geologic period.[29][30][31] Sea levels rose from their Hirnantian low throughout the first half of the Silurian; they subsequently fell throughout the rest of the period, although smaller scale patterns are superimposed on this general trend; fifteen high-stands (periods when sea levels were above the edge of the continental shelf) can be identified, and the highest Silurian sea level was probably around 140 metres (459 ft) higher than the lowest level reached.[26]
During this period, the Earth entered a warm greenhouse phase, supported by high CO2 levels of 4500 ppm, and warm shallow seas covered much of the equatorial land masses.[32] Early in the Silurian, glaciers retreated back into the South Pole until they almost disappeared in the middle of Silurian.[28] Layers of broken shells (called coquina) provide strong evidence of a climate dominated by violent storms generated then as now by warm sea surfaces.[33]
Perturbations
[edit]The climate and carbon cycle appear to be rather unsettled during the Silurian, which had a higher frequency of isotopic excursions (indicative of climate fluctuations) than any other period.[26] The Ireviken event, Mulde event, and Lau event each represent isotopic excursions following a minor mass extinction[34] and associated with rapid sea-level change. Each one leaves a similar signature in the geological record, both geochemically and biologically; pelagic (free-swimming) organisms were particularly hard hit, as were brachiopods, corals, and trilobites, and extinctions rarely occur in a rapid series of fast bursts.[26][31] The climate fluctuations are best explained by a sequence of glaciations, but the lack of tillites in the middle to late Silurian make this explanation problematic.[35]
Flora and fauna
[edit]The Silurian period has been viewed by some palaeontologists as an extended recovery interval following the Late Ordovician mass extinction (LOME), which interrupted the cascading increase in biodiversity that had continuously gone on throughout the Cambrian and most of the Ordovician.[36]
The Silurian was the first period to see megafossils of extensive terrestrial biota in the form of moss-like miniature forests along lakes and streams and networks of large, mycorrhizal nematophytes, heralding the beginning of the Silurian-Devonian Terrestrial Revolution.[12][13][37] However, the land fauna did not have a major impact on the Earth until it diversified in the Devonian.[26]
.jpg)
The first fossil records of vascular plants, that is, land plants with tissues that carry water and food, appeared in the second half of the Silurian Period.[38] The earliest-known representatives of this group are Cooksonia. Most of the sediments containing Cooksonia are marine in nature. Preferred habitats were likely along rivers and streams. Baragwanathia appears to be almost as old, dating to the early Ludlow (420 million years)[needs update?] and has branching stems and needle-like leaves of 10–20 centimetres (3.9–7.9 in). The plant shows a high degree of development in relation to the age of its fossil remains. Fossils of this plant have been recorded in Australia,[39][40] Canada,[41] and China.[42] Eohostimella heathana is an early, probably terrestrial, "plant" known from compression fossils[43] of Early Silurian (Llandovery) age.[44] The chemistry of its fossils is similar to that of fossilised vascular plants, rather than algae.[43]
Fossils that are considered as terrestrial animals are also known from the Silurian. The definitive oldest record of millipede ever known is Kampecaris obanensis and Archidesmus sp. from the late Silurian (425 million years ago) of Kerrera.[45] There are also other millipedes, centipedes, and trigonotarbid arachnoids known from Ludlow (420 million years ago).[45][46][47] Predatory invertebrates would indicate that simple food webs were in place that included non-predatory prey animals. Extrapolating back from Early Devonian biota, Andrew Jeram et al. in 1990[48] suggested a food web based on as-yet-undiscovered detritivores and grazers on micro-organisms.[49] Millipedes from Cowie Formation such as Cowiedesmus and Pneumodesmus were considered as the oldest millipede from the middle Silurian at 428–430 million years ago,[24][50][51] although the age of this formation is later reinterpreted to be from the early Devonian instead by some researchers.[52][53] Regardless, Pneumodesmus is still an important fossil as the oldest definitive evidence of spiracles to breath in the air.[45]

The first bony fish, the Osteichthyes, appeared, represented by the Acanthodians covered with bony scales. Fish reached considerable diversity and developed movable jaws, adapted from the supports of the front two or three gill arches. A diverse fauna of eurypterids (sea scorpions)—some of them several meters in length—prowled the shallow Silurian seas and lakes of North America; many of their fossils have been found in New York state. Brachiopods were abundant and diverse, with the taxonomic composition, ecology, and biodiversity of Silurian brachiopods mirroring Ordovician ones.[54] Brachiopods that survived the LOME developed novel adaptations for environmental stress,[55] and they tended to be endemic to a single palaeoplate in the mass extinction's aftermath, but expanded their range afterwards.[56] The most abundant brachiopods were atrypids and pentamerides;[57] atrypids were the first to recover and rediversify in the Rhuddanian after LOME,[58] while pentameride recovery was delayed until the Aeronian.[57] Bryozoans exhibited significant degrees of endemism to a particular shelf.[59] They also developed symbiotic relationships with cnidarians[60] and stromatolites.[61] Many bivalve fossils have also been found in Silurian deposits,[62] and the first deep-boring bivalves are known from this period.[63] Chitons saw a peak in diversity during the middle of the Silurian.[64] Hederelloids enjoyed significant success in the Silurian, with some developing symbioses with the colonial rugose coral Entelophyllum.[65] The Silurian was a heyday for tentaculitoids,[66] which experienced an evolutionary radiation focused mainly in Baltoscandia,[67] along with an expansion of their geographic range in the Llandovery and Wenlock.[68] Trilobites started to recover in the Rhuddanian,[69] and they continued to enjoy success in the Silurian as they had in the Ordovician despite their reduction in clade diversity as a result of LOME.[70] The Early Silurian was a chaotic time of turnover for crinoids as they rediversified after LOME.[71] Members of Flexibilia, which were minimally impacted by LOME, took on an increasing ecological prominence in Silurian seas.[72] Monobathrid camerates, like flexibles, diversified in the Llandovery, whereas cyathocrinids and dendrocrinids diversified later in the Silurian.[73] Scyphocrinoid loboliths suddenly appeared in the terminal Silurian, shortly before the Silurian-Devonian boundary, and disappeared as abruptly as they appeared very shortly after their first appearance.[74] Endobiotic symbionts were common in the corals and stromatoporoids.[75][76] Rugose corals especially were colonised and encrusted by a diverse range of epibionts,[77] including certain hederelloids as aforementioned.[65] Photosymbiotic scleractinians made their first appearance during the Middle Silurian.[78] Reef abundance was patchy; sometimes, fossils are frequent, but at other points, are virtually absent from the rock record.[26]
-
 Cooksonia, the earliest vascular plant, middle Silurian
Cooksonia, the earliest vascular plant, middle Silurian -
 Silurian sea bed fossils collected from Wren's Nest Nature Reserve, Dudley UK
Silurian sea bed fossils collected from Wren's Nest Nature Reserve, Dudley UK -

-
 Silurian sea bed fossils collected from Wren's Nest Nature Reserve, Dudley UK
Silurian sea bed fossils collected from Wren's Nest Nature Reserve, Dudley UK -
 Eurypterus, a common Upper Silurian eurypterid
Eurypterus, a common Upper Silurian eurypterid -
 Pterygotus was a giant eurypterid that had a nearly cosmopolitan distribution (reconstruction shown here is Devonian species P. anglicus)
Pterygotus was a giant eurypterid that had a nearly cosmopolitan distribution (reconstruction shown here is Devonian species P. anglicus) -
 Trilobites were still diverse and common in the Silurian. Fossils of Calymene celebra are extremely abundant in parts of central US.
Trilobites were still diverse and common in the Silurian. Fossils of Calymene celebra are extremely abundant in parts of central US. -
 Halysites was a Tabulate coral, an extinct group that lived through the Paleozoic
Halysites was a Tabulate coral, an extinct group that lived through the Paleozoic -
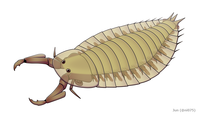 Parioscorpio was an enigmatic arthropod from the Silurian of Wisconsin
Parioscorpio was an enigmatic arthropod from the Silurian of Wisconsin -
 A Dalmanites limulurus specimen from Silurian strata of New York
A Dalmanites limulurus specimen from Silurian strata of New York -
 A rock containing several geodized pentamerid brachiopods from strata in Indiana
A rock containing several geodized pentamerid brachiopods from strata in Indiana -

-

-
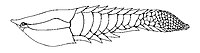
-
 Tujiaaspis is a galeaspid agnathan from the early Silurian (Telychian) of China, showing origin of paired fins
Tujiaaspis is a galeaspid agnathan from the early Silurian (Telychian) of China, showing origin of paired fins -

-
 Fanjingshania, climatiid spiny shark from the lower Silurian (Aeronian) described from disarticulated dermoskeletal elements
Fanjingshania, climatiid spiny shark from the lower Silurian (Aeronian) described from disarticulated dermoskeletal elements -
![Shenacanthus vermiformis[69] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China](//upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Shenacanthus_vermiformis.jpg/154px-Shenacanthus_vermiformis.jpg) Shenacanthus[80] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China
Shenacanthus[80] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China -
![Xiushanosteus[80] is the oldest known placoderm from the early Silurian (Telychian) of China](//upload.wikimedia.org/wikipedia/commons/thumb/c/c5/Xiushanosteus.jpg/200px-Xiushanosteus.jpg) Xiushanosteus[80] is the oldest known placoderm from the early Silurian (Telychian) of China
Xiushanosteus[80] is the oldest known placoderm from the early Silurian (Telychian) of China -
 Entelognathus primordialis was a Placoderm fish from the late Silurian
Entelognathus primordialis was a Placoderm fish from the late Silurian










_1.jpg)





![Shenacanthus vermiformis[69] is jawed stem-chondrichthyan genus from the early Silurian (Telychian) of China](/wiki/File:Shenacanthus_vermiformis.jpg)
![Xiushanosteus[80] is the oldest known placoderm from the early Silurian (Telychian) of China](/wiki/File:Xiushanosteus.jpg)

{kind=link}
{kind=link}
See also
[edit]Notes
[edit]- ^ Jeppsson, L.; Calner, M. (2007). "The Silurian Mulde Event and a scenario for secundo—secundo events". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 93 (02): 135–154. doi:10.1017/S0263593300000377.
- ^ Munnecke, A.; Samtleben, C.; Bickert, T. (2003). "The Ireviken Event in the lower Silurian of Gotland, Sweden-relation to similar Palaeozoic and Proterozoic events". Palaeogeography, Palaeoclimatology, Palaeoecology. 195 (1): 99–124. doi:10.1016/S0031-0182(03)00304-3.
- ^ "International Chronostratigraphic Chart" (PDF). International Commission on Stratigraphy. September 2023. Retrieved December 16, 2024.
- ^ Lucas, Sepncer (6 November 2018). "The GSSP Method of Chronostratigraphy: A Critical Review". Frontiers in Earth Science. 6: 191. Bibcode:2018FrEaS...6..191L. doi:10.3389/feart.2018.00191.
- ^ Holland, C. (June 1985). "Series and Stages of the Silurian System" (PDF). Episodes. 8 (2): 101–103. doi:10.18814/epiiugs/1985/v8i2/005. Retrieved 11 December 2020.
- ^ Chlupáč, Ivo; Hladil, Jindrich (January 2000). "The global stratotype section and point of the Silurian-Devonian boundary". CFS Courier Forschungsinstitut Senckenberg. Retrieved 7 December 2020.
- ^ Haq, B. U.; Schutter, SR (2008). "A Chronology of Paleozoic Sea-Level Changes". Science. 322 (5898): 64–68. Bibcode:2008Sci...322...64H. doi:10.1126/science.1161648. PMID 18832639. S2CID 206514545.
- ^ Wells, John (3 April 2008). Longman Pronunciation Dictionary (3rd ed.). Pearson Longman. ISBN 978-1-4058-8118-0.
- ^ "Silurian". Dictionary.com Unabridged (Online). n.d.
- ^ "Silurian". CollinsDictionary.com. HarperCollins.
- ^ "International Chronostratigraphic Chart v.2015/01" (PDF). International Commission on Stratigraphy. January 2015.
- ^ a b Capel, Elliot; Cleal, Christopher J.; Xue, Jinzhuang; Monnet, Claude; Servais, Thomas; Cascales-Miñana, Borja (August 2022). "The Silurian–Devonian terrestrial revolution: Diversity patterns and sampling bias of the vascular plant macrofossil record". Earth-Science Reviews. 231: 104085. Bibcode:2022ESRv..23104085C. doi:10.1016/j.earscirev.2022.104085. hdl:20.500.12210/76731. S2CID 249616013.
- ^ a b Xue, Jinzhuang; Huang, Pu; Wang, Deming; Xiong, Conghui; Liu, Le; Basinger, James F. (May 2018). "Silurian-Devonian terrestrial revolution in South China: Taxonomy, diversity, and character evolution of vascular plants in a paleogeographically isolated, low-latitude region". Earth-Science Reviews. 180: 92–125. Bibcode:2018ESRv..180...92X. doi:10.1016/j.earscirev.2018.03.004. Retrieved 8 December 2022.
- ^ Lutzoni, François; Nowak, Michael D.; Alfaro, Michael E.; Reeb, Valérie; Miadlikowska, Jolanta; Krug, Michael; Arnold, A. Elizabeth; Lewis, Louise A.; Swofford, David L.; Hibbett, David; Hilu, Khidir; James, Timothy Y.; Quandt, Dietmar; Magallón, Susana (21 December 2018). "Contemporaneous radiations of fungi and plants linked to symbiosis". Nature Communications. 9 (1): 5451. Bibcode:2018NatCo...9.5451L. doi:10.1038/s41467-018-07849-9. PMC 6303338. PMID 30575731. S2CID 56645104.
- ^ Garwood, Russell J.; Edgecombe, Gregory D. (September 2011). "Early Terrestrial Animals, Evolution, and Uncertainty". Evolution: Education and Outreach. 4 (3): 489–501. doi:10.1007/s12052-011-0357-y.
- ^ Brazeau, M. D.; Friedman, M. (2015). "The origin and early phylogenetic history of jawed vertebrates". Nature. 520 (7548): 490–497. Bibcode:2015Natur.520..490B. doi:10.1038/nature14438. PMC 4648279. PMID 25903631.
- ^ See:
- Murchison, Roderick Impey (1835). "On the Silurian system of rocks". Philosophical Magazine. 3rd series. 7 (37): 46–52. doi:10.1080/14786443508648654. From p. 48: " … I venture to suggest, that as the great mass of rocks in question, trending from south-west to north-east, traverses the kingdom of our ancestors the Silures, the term "Silurian system" should be adopted … "
- Wilmarth, Mary Grace (1925). Bulletin 769: The Geologic Time Classification of the United States Geological Survey Compared With Other Classifications, accompanied by the original definitions of era, period and epoch terms. Washington, D.C., U.S.: U.S. Government Printing Office. p. 80.
- ^ Murchison, Roderick (1835). "On the Silurian System of Rocks". The London and Edinburgh Philosophical Magazine and Journal of Science. Third Series, Vol. 7: 46–52 – via Biodiversity Heritage Library.
- ^ Sedgwick; Murchison, R.I. (1835). "On the Silurian and Cambrian systems, exhibiting the order in which the older sedimentary strata succeed each other in England and Wales". Report of the Fifth Meeting of the British Association for the Advancement of Science. § Notices and Abstracts of Miscellaneous Communications to the Sections. 5: 59–61.
- ^ Lapworth, Charles (1879). "On the tripartite classification of the Lower Palaeozoic rocks". Geological Magazine. 2nd series. 6 (1): 1–15. Bibcode:1879GeoM....6....1L. doi:10.1017/s0016756800156560. S2CID 129165105. From pp. 13–14: "North Wales itself – at all events the whole of the great Bala district where Sedgwick first worked out the physical succession among the rocks of the intermediate or so-called Upper Cambrian or Lower Silurian system; and in all probability much of the Shelve and the Caradoc area, whence Murchison first published its distinctive fossils – lay within the territory of the Ordovices; … Here, then, have we the hint for the appropriate title for the central system of the Lower Palaeozoics. It should be called the Ordovician System, after this old British tribe."
- ^ The Gotlandian system was proposed in 1893 by the French geologist Albert Auguste Cochon de Lapparent (1839–1908): Lapparent, A. de (1893). Traité de Géologie (in French). Vol. 2 (3rd ed.). Paris, France: F. Savy. p. 748. From p. 748: "D'accord avec ces divisions, on distingue communément dans le silurien trois étages: l'étage inférieur ou cambrien (1); l'étage moyen ou ordovicien (2); l'étage supérieur ou gothlandien (3)." (In agreement with these divisions, one generally distinguishes, within the Silurian, three stages: the lower stage or Cambrian [1]; the middle stage or Ordovician [2]; the upper stage or Gotlandian [3].)
- ^ Barrande, Joachim (1852). Systême silurien du centre de la Bohême (in French). Paris, France and Prague, (Czech Republic): (Self-published). pp. ix–x.
- ^ Forbes, Edward (1854). "Anniversary Address of the President". Quarterly Journal of the Geological Society of London. 10: xxii–lxxxi. See p. xxxiv.
- ^ a b Paul Selden & Helen Read (2008). "The oldest land animals: Silurian millipedes from Scotland" (PDF). Bulletin of the British Myriapod & Isopod Group. 23: 36–37.
- ^ Manda, Štěpán; Frýda, Jiří (2010). "Silurian-Devonian boundary events and their influence on cephalopod evolution: evolutionary significance of cephalopod egg size during mass extinctions". Bulletin of Geosciences. 85 (3): 513–40. doi:10.3140/bull.geosci.1174.
- ^ a b c d e f g Munnecke, Axel; Calner, Mikael; Harper, David A.T.; Servais, Thomas (2010). "Ordovician and Silurian sea–water chemistry, sea level, and climate: A synopsis". Palaeogeography, Palaeoclimatology, Palaeoecology. 296 (3–4): 389–413. Bibcode:2010PPP...296..389M. doi:10.1016/j.palaeo.2010.08.001.
- ^ Yan, Guanzhou; Lehnert, Oliver; Männik, Peep; Calner, Mikael; Luan, Xiaocong; Gong, Fanyi; Li, Lixia; Wei, Xin; Wang, Guangxu; Zhan, Renbin; Wu, Rongchang (15 November 2022). "The record of early Silurian climate changes from South China and Baltica based on integrated conodont biostratigraphy and isotope chemostratigraphy". Palaeogeography, Palaeoclimatology, Palaeoecology. 606: 111245. Bibcode:2022PPP...60611245Y. doi:10.1016/j.palaeo.2022.111245. S2CID 252504361. Retrieved 8 December 2022.
- ^ a b Gambacorta, G.; Menichetti, E.; Trincianti, E.; Torricelli, S. (March 2019). "The Silurian climatic transition recorded in the epicontinental Baltica Sea". Palaeogeography, Palaeoclimatology, Palaeoecology. 517: 16–29. Bibcode:2019PPP...517...16G. doi:10.1016/j.palaeo.2018.12.016. S2CID 135118794.
- ^ Young, Set A.; Benayoun, Emily; Kozik, Nevin P.; Hints, Olle; Martma, Tõnu; Bergström, Stig M.; Owens, Jeremy D. (15 September 2020). "Marine redox variability from Baltica during extinction events in the latest Ordovician–early Silurian". Palaeogeography, Palaeoclimatology, Palaeoecology. 554: 109792. Bibcode:2020PPP...55409792Y. doi:10.1016/j.palaeo.2020.109792. S2CID 218930512.
- ^ Sproson, Adam D. (15 February 2020). "Pacing of the latest Ordovician and Silurian carbon cycle by a ~4.5 Myr orbital cycle". Palaeogeography, Palaeoclimatology, Palaeoecology. 540: 109543. Bibcode:2020PPP...54009543S. doi:10.1016/j.palaeo.2019.109543. S2CID 213445668. Retrieved 8 December 2022.
- ^ a b Trotter, Julie A.; Williams, Ian S.; Barnes, Christopher R.; Männik, Peep; Simpson, Andrew (February 2016). "New conodont δ18O records of Silurian climate change: Implications for environmental and biological events". Palaeogeography, Palaeoclimatology, Palaeoecology. 443: 34–48. Bibcode:2016PPP...443...34T. doi:10.1016/j.palaeo.2015.11.011.
- ^ Chaloner, William G. (2003). "The role of carbon dioxide in plant evolution". Evolution on Planet Earth: 65–83. doi:10.1016/B978-012598655-7/50032-X. ISBN 9780125986557.
- ^ Nealon, T.; Williams, D. Michael (30 April 2007). "Storm-influenced shelf deposits from the silurian of Western Ireland: A reinterpretation of deep basin sediments". Geological Journal. 23 (4): 311–320. doi:10.1002/gj.3350230403.
- ^ Samtleben, C.; Munnecke, A.; Bickert, T. (2000). "Development of facies and C/O-isotopes in transects through the Ludlow of Gotland: Evidence for global and local influences on a shallow-marine environment". Facies. 43 (1): 1–38. Bibcode:2000Faci...43....1S. doi:10.1007/BF02536983. S2CID 130640332.
- ^ Calner, Mikael (2008). "Silurian global events – at the tipping point of climate change". Mass Extinction. pp. 21–57. doi:10.1007/978-3-540-75916-4_4. ISBN 978-3-540-75915-7.
- ^ Rasmussen, Christian M. Ø.; Kröger, Björn; Nielsen, Morten L.; Colmenar, Jorge (9 April 2019). "Cascading trend of Early Paleozoic marine radiations paused by Late Ordovician extinctions". Proceedings of the National Academy of Sciences of the United States of America. 116 (15): 7207–7213. Bibcode:2019PNAS..116.7207R. doi:10.1073/pnas.1821123116. PMC 6462056. PMID 30910963.
- ^ Retallack, Gregory J. (June 2022). "Ordovician-Devonian lichen canopies before evolution of woody trees". Gondwana Research. 106: 211–223. Bibcode:2022GondR.106..211R. doi:10.1016/j.gr.2022.01.010. S2CID 246320087. Retrieved 23 November 2022.
- ^ Rittner, Don (2009). Encyclopedia of Biology. Infobase Publishing. p. 338. ISBN 9781438109992.
- ^ Rickards, R. B. (1 March 2000). "The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia". Geological Magazine. 137 (2): 207–209. Bibcode:2000GeoM..137..207R. doi:10.1017/S0016756800003800. S2CID 131287538. Retrieved 8 December 2022.
- ^ Lang, W.H.; Cookson, I.C. (1935). "On a flora, including vascular land plants, associated with Monograptus, in rocks of Silurian age, from Victoria, Australia". Philosophical Transactions of the Royal Society of London B. 224 (517): 421–449. Bibcode:1935RSPTB.224..421L. doi:10.1098/rstb.1935.0004.
- ^ Hueber, F.M. (1983). "A new species of Baragwanathia from the Sextant Formation (Emsian) Northern Ontario, Canada". Botanical Journal of the Linnean Society. 86 (1–2): 57–79. doi:10.1111/j.1095-8339.1983.tb00717.x.
- ^ Bora, Lily (2010). Principles of Paleobotany. Mittal Publications. pp. 36–37.
- ^ a b Niklas, Karl J. (1976). "Chemical Examinations of Some Non-Vascular Paleozoic Plants". Brittonia. 28 (1): 113–137. Bibcode:1976Britt..28..113N. doi:10.2307/2805564. JSTOR 2805564. S2CID 21794174.
- ^ Edwards, D. & Wellman, C. (2001), "Embryophytes on Land: The Ordovician to Lochkovian (Lower Devonian) Record", in Gensel, P. & Edwards, D. (eds.), Plants Invade the Land : Evolutionary and Environmental Perspectives, New York: Columbia University Press, pp. 3–28, ISBN 978-0-231-11161-4, p. 4
- ^ a b c Brookfield, M. E.; Catlos, E. J.; Suarez, S. E. (2021-10-03). "Myriapod divergence times differ between molecular clock and fossil evidence: U/Pb zircon ages of the earliest fossil millipede-bearing sediments and their significance". Historical Biology. 33 (10): 2014–2018. Bibcode:2021HBio...33.2014B. doi:10.1080/08912963.2020.1762593. ISSN 0891-2963. S2CID 238220137.
- ^ Dunlop, Jason A. (1996). "A trigonotarbid arachnid from the Upper Silurian of Shropshire". Palaeontology. 39: 605–614. ISSN 0031-0239.
- ^ Shear, William A.; Jeram, Andrew J.; Selden, Paul (1998). Centiped legs (Arthropoda, Chilopoda, Scutigeromorpha) from the Silurian and Devonian of Britain and the Devonian of North America. New York, NY: American Museum of Natural History.
- ^ Jeram, Andrew J.; Selden, Paul A.; Edwards, Dianne (1990). "Land Animals in the Silurian: Arachnids and Myriapods from Shropshire, England". Science. 250 (4981): 658–61. Bibcode:1990Sci...250..658J. doi:10.1126/science.250.4981.658. PMID 17810866.
- ^ DiMichele, William A; Hook, Robert W (1992). "The Silurian". In Behrensmeyer, Anna K. (ed.). Terrestrial Ecosystems Through Time: Evolutionary Paleoecology of Terrestrial Plants and Animals. University of Chicago Press. pp. 207–10. ISBN 978-0-226-04155-1.
- ^ Wilson, Heather M.; Anderson, Lyall I. (2004). "Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland". Journal of Paleontology. 78 (1): 169–184. Bibcode:2004JPal...78..169W. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2. ISSN 0022-3360. S2CID 131201588.
- ^ Wellman, C.H.; Lopes, G.; McKellar, Z.; Hartley, A. (2023). "Age of the basal 'Lower Old Red Sandstone' Stonehaven Group of Scotland: The oldest reported air-breathing land animal is Silurian (late Wenlock) in age". Journal of the Geological Society. 181. The Geological Society of London. doi:10.1144/jgs2023-138. hdl:2164/22754. ISSN 0016-7649.
- ^ Suarez, Stephanie E.; Brookfield, Michael E.; Catlos, Elizabeth J.; Stöckli, Daniel F. (2017-06-28). "A U-Pb zircon age constraint on the oldest-recorded air-breathing land animal". PLOS ONE. 12 (6): e0179262. Bibcode:2017PLoSO..1279262S. doi:10.1371/journal.pone.0179262. ISSN 1932-6203. PMC 5489152. PMID 28658320.
- ^ Brookfield, M. E.; Catlos, E. J.; Garza, H. (2024-07-07). "The oldest 'millipede'-plant association? Age, paleoenvironments and sources of the Silurian lake sediments at Kerrera, Argyll and Bute, Scotland". Historical Biology: 1–13. doi:10.1080/08912963.2024.2367554. ISSN 0891-2963.
- ^ Rong, Jia-yu; Shen, Shu-zhong (1 December 2002). "Comparative analysis of the end-Permian and end-Ordovician brachiopod mass extinctions and survivals in South China". Palaeogeography, Palaeoclimatology, Palaeoecology. 188 (1–2): 25–38. Bibcode:2002PPP...188...25R. doi:10.1016/S0031-0182(02)00507-2. Retrieved 16 April 2023.
- ^ Rong, Jiayu; Zhan, Renbin (October 1999). "Chief sources of brachiopod recovery from the end Ordovician mass extinction with special references to progenitors". Science in China Series D: Earth Sciences. 42 (5): 553–560. Bibcode:1999ScChD..42..553R. doi:10.1007/BF02875250. S2CID 129323463. Retrieved 16 April 2023.
- ^ Huang, Bing; Rong, Jiayu; Cocks, L. Robin M. (1 February 2012). "Global palaeobiogeographical patterns in brachiopods from survival to recovery after the end-Ordovician mass extinction". Palaeogeography, Palaeoclimatology, Palaeoecology. 317–318: 196–205. Bibcode:2012PPP...317..196H. doi:10.1016/j.palaeo.2012.01.009. Retrieved 16 April 2023.
- ^ a b Huang, Bing; Jin, Jisuo; Rong, Jia-Yu (15 March 2018). "Post-extinction diversification patterns of brachiopods in the early–middle Llandovery, Silurian". Palaeogeography, Palaeoclimatology, Palaeoecology. 493: 11–19. Bibcode:2018PPP...493...11H. doi:10.1016/j.palaeo.2017.12.025. Retrieved 23 November 2022.
- ^ Jia-Yu, Rong; Harper, David A. T. (31 January 2000). "Brachiopod survival and recovery from the latest Ordovician mass extinctions in South China". Geological Journal. 34 (4): 321–348. doi:10.1002/(SICI)1099-1034(199911/12)34:4<321::AID-GJ809>3.0.CO;2-I. Retrieved 16 April 2023.
- ^ Anstey, Robert L.; Pachut, Joseph F.; Tuckey, Michael E. (8 April 2016). "Patterns of bryozoan endemism through the Ordovician-Silurian transition". Paleobiology. 29 (3): 305–328. doi:10.1666/0094-8373(2003)0292.0.CO;2 (inactive 1 November 2024). Retrieved 16 April 2023.
{{cite journal}}: CS1 maint: DOI inactive as of November 2024 (link) - ^ Zapalski, Mikołaj K.; Vinn, Olev; Toom, Ursula; Ernst, Andrej; Wilson, Mark A. (16 September 2022). "Bryozoan–cnidarian mutualism triggered a new strategy for greater resource exploitation as early as the Late Silurian". Scientific Reports. 12 (1): 15556. Bibcode:2022NatSR..1215556Z. doi:10.1038/s41598-022-19955-2. PMC 9481587. PMID 36114227.
- ^ Claussen, Anna Lene; Munnecke, Axel; Ernst, Andrej (2 March 2021). "Bryozoan-rich stromatolites (bryostromatolites) from the Silurian of Gotland and their relation to climate-related perturbations of the global carbon cycle". Sedimentology. 69 (1): 162–198. doi:10.1111/sed.12863. S2CID 233820331.
- ^ Liljedahl, Louis (January 1985). "Ecological aspects of a silicified bivalve fauna from the Silurian of Gotland". Lethaia. 18 (1): 53–66. Bibcode:1985Letha..18...53L. doi:10.1111/j.1502-3931.1985.tb00684.x. Retrieved 16 April 2023.
- ^ Claussen, Anna Lene; Munnecke, Axel; Wilson, Mark A.; Oswald, Irina (22 April 2019). "The oldest deep-boring bivalves? Evidence from the Silurian of Gotland (Sweden)". Facies. 65 (3): 26. Bibcode:2019Faci...65...26C. doi:10.1007/s10347-019-0570-7. S2CID 149500698. Retrieved 16 April 2023.
- ^ Cherns, Lesley (2 January 2007). "Early Palaeozoic diversification of chitons (Polyplacophora, Mollusca) based on new data from the Silurian of Gotland, Sweden". Lethaia. 37 (4): 445–456. doi:10.1080/00241160410002180. Retrieved 16 April 2023.
- ^ a b Vinn, Olev; Wilson, Mark A.; Madison, Anna; Kazantseva, Elizaveta; Toom, Ursula (26 July 2022). "First Symbiotic Association Between Hederelloids and Rugose Corals (Latest Silurian of Saaremaa, Estonia)". PALAIOS. 37 (7): 368–373. Bibcode:2022Palai..37..368V. doi:10.2110/palo.2022.005. S2CID 251122025. Retrieved 16 April 2023.
- ^ Vinn, Olev (1 June 2010). "Adaptive strategies in the evolution of encrusting tentaculitoid tubeworms". Palaeogeography, Palaeoclimatology, Palaeoecology. 292 (1–2): 211–221. Bibcode:2010PPP...292..211V. doi:10.1016/j.palaeo.2010.03.046. Retrieved 16 April 2023.
- ^ Wittmer, Jacalyn M.; Miller, Arnold I. (1 December 2011). "Dissecting the global diversity trajectory of an enigmatic group: The paleogeographic history of tentaculitoids". Palaeogeography, Palaeoclimatology, Palaeoecology. 312 (1–2): 54–65. Bibcode:2011PPP...312...54W. doi:10.1016/j.palaeo.2011.09.009. Retrieved 16 April 2023.
- ^ Wei, Fan; Gong, Yiming; Yang, Hao (1 November 2012). "Biogeography, ecology and extinction of Silurian and Devonian tentaculitoids". Palaeogeography, Palaeoclimatology, Palaeoecology. 358–360: 40–50. Bibcode:2012PPP...358...40W. doi:10.1016/j.palaeo.2012.07.019. Retrieved 16 April 2023.
- ^ Wei, Xin; Zhan, Ren-Bin (March 2018). "A late Rhuddanian (early Llandovery, Silurian) trilobite association from South China and its implications". Palaeoworld. 27 (1): 42–52. doi:10.1016/j.palwor.2017.05.008. Retrieved 16 April 2023.
- ^ Adrain, Jonathan M.; Westrop, Stephen R.; Chatterton, Brian D. E.; Ramsköld, Lars (8 February 2016). "Silurian trilobite alpha diversity and the end-Ordovician mass extinction". Paleobiology. 26 (4): 625–646. doi:10.1666/0094-8373(2000)0262.0.CO;2 (inactive 1 November 2024). Retrieved 16 April 2023.
{{cite journal}}: CS1 maint: DOI inactive as of November 2024 (link) - ^ Peters, Shanan E.; Ausich, William I. (8 April 2016). "A sampling-adjusted macroevolutionary history for Ordovician-Early Silurian crinoids". Paleobiology. 34 (1): 104–116. doi:10.1666/07035.1. S2CID 67838459. Retrieved 16 April 2023.
- ^ Wright, David F.; Toom, Ursula (30 August 2017). "New crinoids from the Baltic region (Estonia): fossil tip-dating phylogenetics constrains the origin and Ordovician–Silurian diversification of the Flexibilia (Echinodermata)". Palaeontology. 60 (6): 893–910. Bibcode:2017Palgy..60..893W. doi:10.1111/pala.12324. S2CID 134049045.
- ^ Ausich, William I.; Deline, Bradley (15 November 2012). "Macroevolutionary transition in crinoids following the Late Ordovician extinction event (Ordovician to Early Silurian)". Palaeogeography, Palaeoclimatology, Palaeoecology. 361–362: 38–48. Bibcode:2012PPP...361...38A. doi:10.1016/j.palaeo.2012.07.022. Retrieved 16 April 2023.
- ^ Zong, Rui-wen; Gong, Yi-ming (1 November 2020). "Discovery of scyphocrinoid loboliths in western Junggar, Xinjiang, NW China: Implications for scyphocrinoid paleobiogeography and identification of the Silurian–Devonian boundary". Palaeogeography, Palaeoclimatology, Palaeoecology. 557: 109914. Bibcode:2020PPP...55709914Z. doi:10.1016/j.palaeo.2020.109914. S2CID 224964119. Retrieved 16 April 2023.
- ^ Vinn, O.; wilson, M.A.; Mõtus, M.-A. (2014). "Symbiotic endobiont biofacies in the Silurian of Baltica". Palaeogeography, Palaeoclimatology, Palaeoecology. 404: 24–29. Bibcode:2014PPP...404...24V. doi:10.1016/j.palaeo.2014.03.041. Retrieved 2014-06-11.
- ^ Vinn, O.; Mõtus, M.-A. (2008). "The earliest endosymbiotic mineralized tubeworms from the Silurian of Podolia, Ukraine". Journal of Paleontology. 82 (2): 409–414. Bibcode:2008JPal...82..409V. doi:10.1666/07-056.1. S2CID 131651974. Retrieved 2014-06-11.
- ^ Zatoń, Michał; Wrzołek, Tomasz (15 October 2020). "Colonization of rugose corals by diverse epibionts: dominance and syn vivo encrustation in a Middle Devonian (Givetian) soft-bottom habitat of the Holy Cross Mountains, Poland". Palaeogeography, Palaeoclimatology, Palaeoecology. 556: 109899. Bibcode:2020PPP...55609899Z. doi:10.1016/j.palaeo.2020.109899. S2CID 224869051. Retrieved 16 April 2023.
- ^ Zapalski, Mikołaj K. (22 January 2014). "Evidence of photosymbiosis in Palaeozoic tabulate corals". Proceedings of the Royal Society B: Biological Sciences. 281 (1775): 20132663. doi:10.1098/rspb.2013.2663. ISSN 0962-8452. PMC 3866410. PMID 24307674.
- ^ Andreev, Plamen S.; Sansom, Ivan J.; Li, Qiang; Zhao, Wenjin; Wang, Jianhua; Wang, Chun-Chieh; Peng, Lijian; Jia, Liantao; Qiao, Tuo; Zhu, Min (September 2022). "The oldest gnathostome teeth". Nature. 609 (7929): 964–968. Bibcode:2022Natur.609..964A. doi:10.1038/s41586-022-05166-2. ISSN 1476-4687. PMID 36171375. S2CID 252569771.
- ^ a b Zhu, You-an; Li, Qiang; Lu, Jing; Chen, Yang; Wang, Jianhua; Gai, Zhikun; Zhao, Wenjin; Wei, Guangbiao; Yu, Yilun; Ahlberg, Per E.; Zhu, Min (2022). "The oldest complete jawed vertebrates from the early Silurian of China". Nature. 609 (7929): 954–958. Bibcode:2022Natur.609..954Z. doi:10.1038/s41586-022-05136-8. ISSN 1476-4687. PMID 36171378. S2CID 252569910.
References
[edit]- Emiliani, Cesare. (1992). Planet Earth : Cosmology, Geology, & the Evolution of Life & the Environment. Cambridge University Press. (Paperback Edition ISBN 0-521-40949-7)
- Mikulic, DG, DEG Briggs, and J Kluessendorf. 1985. A new exceptionally preserved biota from the Lower Silurian of Wisconsin, USA. Philosophical Transactions of the Royal Society of London, 311B:75-86.
- Moore, RA; Briggs, DEG; Braddy, SJ; Anderson, LI; Mikulic, DG; Kluessendorf, J (2005). "A new synziphosurine (Chelicerata: Xiphosura) from the Late Llandovery (Silurian) Waukesha Lagerstatte, Wisconsin, USA". Journal of Paleontology. 79 (2): 242–250. Bibcode:2005JPal...79..242M. doi:10.1666/0022-3360(2005)079<0242:anscxf>2.0.co;2. S2CID 56570105.
External links
[edit]- Ogg, Jim (June 2004), Overview of Global Boundary Stratotype Sections and Points (GSSP's)
- Palaeos: Silurian
- UCMP Berkeley: The Silurian
- Paleoportal: Silurian strata in U.S., state by state Archived 2021-03-15 at the Wayback Machine
- USGS:Silurian and Devonian Rocks (U.S.)
- "International Commission on Stratigraphy (ICS)". Geologic Time Scale 2004. Retrieved September 19, 2005.
- Examples of Silurian Fossils
- GeoWhen Database for the Silurian
- Silurian (Chronostratography scale)